How Do You Know if an Element Is Limiting in Open Ocean? Graph
What is Ocean Productivity?
Ocean productivity largely refers to the production of organic affair by "phytoplankton," plants suspended in the body of water, most of which are single-celled. Phytoplankton are "photoautotrophs," harvesting lite to convert inorganic to organic carbon, and they supply this organic carbon to diverse "heterotrophs," organisms that obtain their energy solely from the respiration of organic matter. Open bounding main heterotrophs include bacteria too equally more complex unmarried- and multi-celled "zooplankton" (floating animals), "nekton" (swimming organisms, including fish and marine mammals), and the "benthos" (the seafloor community of organisms).
The many nested cycles of carbon associated with ocean productivity are revealed by the following definitions (Bender et al. 1987) (Figure 1). "Gross primary product" (GPP) refers to the total charge per unit of organic carbon production past autotrophs, while "respiration" refers to the free energy-yielding oxidation of organic carbon back to carbon dioxide. "Net primary product" (NPP) is GPP minus the autotrophs' own rate of respiration; information technology is thus the charge per unit at which the total metabolism of phytoplankton produces biomass. "Secondary production" (SP) typically refers to the growth charge per unit of heterotrophic biomass. Only a minor fraction of the organic matter ingested by heterotrophic organisms is used to grow, the majority beingness respired back to dissolved inorganic carbon and nutrients that tin can be reused by autotrophs. Therefore, SP in the bounding main is small in comparing to NPP. Fisheries rely on SP; thus they depend on both NPP and the efficiency with which organic affair is transferred up the foodweb (i.east., the SP/NPP ratio). "Cyberspace ecosystem production" (NEP) is GPP minus the respiration by all organisms in the ecosystem. The value of NEP depends on the boundaries defined for the ecosystem. If one considers the sunlit surface ocean down to the i% lite level (the "euphotic zone") over the course of an entire year, then NEP is equivalent to the particulate organic carbon sinking into the night ocean interior plus the dissolved organic carbon being circulated out of the euphotic zone. In this case, NEP is also often referred to as "export production" (or "new product" (Dugdale & Goering 1967), as discussed below). In contrast, the NEP for the entire body of water, including its shallow sediments, is roughly equivalent to the slow burial of organic matter in the sediments minus the rate of organic matter inbound from the continents.
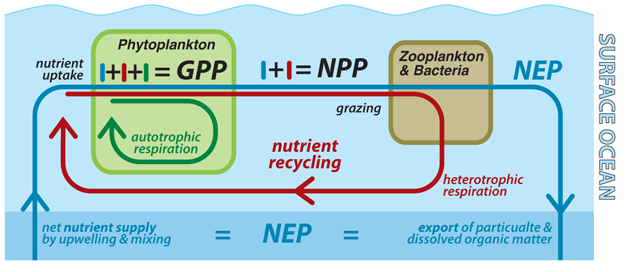
Figure 1
Productivity in the surface ocean, the definitions used to describe it, and its connections to food cycling. The bluish bike for "net ecosystem production" (NEP) (i.e. "new" or "export" product) encompasses the "new" nutrient supply from the ocean interior, its uptake by autotrophic phytoplankton growth, packaging into large particles by heterotrophic grazing organisms, and sinking of organic matter out of the surface bounding main. The red wheel illustrates the fate of the majority of organic matter produced in the surface body of water, which is to exist respired by heterotrophic organisms to meet their energy requirements, thereby releasing the nutrients dorsum into the surface water where they can be taken up past phytoplankton once again to fuel "regenerated product." The green wheel represents the internal respiration of phytoplankton themselves, that is, their ain utilise of the products of photosynthesis for purposes other than growth. These nested cycles combine to yield (ane) "gross principal production" (GPP) representing the gross photosynthesis and (2) "internet master production" (NPP) that represents phytoplankton biomass production that forms the basis of the food spider web plus a much smaller rate of organic matter consign from the surface. While the new nutrient supply and consign production are ultimately linked past mass balance, at that place may exist imbalances on small scales of space and fourth dimension, assuasive for brief accumulations of biomass.
There are no accumulations of living biomass in the marine environment that compare with the forests and grasslands on land (Sarmiento & Bender 1994). Even so, bounding main biological science is responsible for the storage of more carbon away from the atmosphere than is the terrestrial biosphere (Broecker 1982). This is achieved by the sinking of organic matter out of the surface ocean and into the ocean interior before information technology is returned to dissolved inorganic carbon and dissolved nutrients by bacterial decomposition. Oceanographers often refer to this process as the "biological pump," equally it pumps carbon dioxide (COii) out of the surface ocean and temper and into the voluminous deep ocean (Volk & Hoffert 1985).
Only a fraction of the organic matter produced in the surface bounding main has the fate of being exported to the deep ocean. Of the organic matter produced by phytoplankton (NPP), most is respired back to dissolved inorganic forms within the surface ocean and thus recycled for use by phytoplankton (Eppley & Peterson 1979) (Figure one). Most phytoplankton cells are likewise small to sink individually, and so sinking occurs simply one time they aggregate into larger particles or are packaged into "fecal pellets" past zooplankton. The remains of zooplankton are besides adequately large to sink. While sinking is a relatively rare fate for any given particle in the surface ocean, biomass and organic thing do non accrue in the surface bounding main, so consign of organic thing by sinking is the ultimate fate for all of the nutrients that enter into the surface sea in dissolved form — with the exceptions that (1) dissolved nutrients tin can be returned unused to the interior past the apportionment in some polar regions (come across below), and (2) circulation also carries dissolved organic thing from the surface ocean into the interior, a significant procedure (Hansell et al. 2009) that we will not accost farther. As organic thing settles through the bounding main interior and onto the seafloor, it is about entirely decomposed back to dissolved chemicals (Emerson & Hedges 2003, Martin et al. 1987). This high efficiency of decomposition is due to the fact that the organisms conveying out the decomposition rely upon information technology every bit their sole source of chemical energy; in most of the open ocean, the heterotrophs only go out behind the organic matter that is too chemically resistant for information technology to exist worth the investment to decompose. On the whole, but a tiny fraction (typically much less than i%) of the organic carbon from NPP in the euphotic zone survives to exist buried in deep sea sediments.
Productivity in littoral ecosystems is often singled-out from that of the open ocean. Forth the coasts, the seafloor is shallow, and sunlight tin sometimes penetrate all the way through the water column to the bottom, thus enabling bottom-dwelling ("benthic") organisms to photosynthesize. Furthermore, sinking organic affair isintercepted by the seabed, where it supports thriving benthic faunal communities, in the process being recycled dorsum to dissolved nutrients that are so immediately bachelor for principal product. The proximity to state and its nutrient sources, the interception of sinking organic affair by the shallow seafloor, and the propensity for coastal upwelling all result in highly productive ecosystems. Here, nosotros mainly address the productivity of the vast open ocean; still, many of the same concepts, albeit in modified form, apply to coastal systems.
What Does Ocean Productivity Need?
Phytoplankton require a suite of chemicals, and those with the potential to be scarce in surface waters are typically identified as "nutrients." Calcium is an example of an element that is rapidly alloyed by some plankton (for production of calcium carbonate "hard parts") but is not typically considered a nutrient because of its uniformly high concentration in seawater. Dissolved inorganic carbon, which is the feedstock for organic carbon product by photosynthesis, is also abundant and so is not typically listed among the nutrients. However, its acidic grade dissolved CO2 is ofttimes at adequately depression concentrations to affect the growth of at least some phytoplankton.
Broadly important nutrients include nitrogen (North), phosphorus (P), iron (Fe), and silicon (Si). There appear to be relatively uniform requirements for N and P among phytoplankton. In the early 1900s, oceanographer Alfred Redfield found that plankton build their biomass with C:N:P stoichiometric ratios of ~106:sixteen:i, to which we now refer every bit the Redfield ratios (Redfield 1958). As Redfield noted, the dissolved N:P in the deep ocean is close to the 16:1 ratio of plankton biomass, and we will argue below that plankton impose this ratio on the deep, non vice versa. Iron is institute in biomass simply in trace amounts, but it is used for diverse essential purposes in organisms, and it has become articulate over the concluding 25 years that iron's scarcity oftentimes limits or affects productivity in the open bounding main, especially those regions where high-Due north and -P deep water is brought rapidly to the surface (Martin & Fitzwater 1988). Enquiry is ongoing to understand the office of other trace elements in productivity (Morel et al. 2003). Silicon is a nutrient only for specific plankton taxa-diatoms (autotrophic phytoplankton), silicoflaggellates, and radiolaria (heterotrophic zooplankton) — which use it to make opal difficult parts. Withal, the typical authorisation of diatoms in Si-bearing waters, and the tendency of diatom-associated organic affair to sink out of the surface ocean, brand Si availability a major factor in the broader environmental and biogeochemistry of surface waters.
Sunlight is the ultimate energy source — directly or indirectly — for almost all life on Earth, including in the deep ocean. Still, light is absorbed and scattered such that very lilliputian of it penetrates below a depth of ~80 thou (every bit deep as 150 m in the to the lowest degree productive subtropical regions, just as shallow as 10 grand in highly productive and coastal regions) (Figure 2). Thus, photosynthesis is largely restricted to the upper calorie-free-penetrated skin of the ocean. Moreover, across most of the ocean's area, including the tropics, subtropics, and the temperate zone, the absorption of sunlight causes surface water to be much warmer than the underlying deep ocean, the latter being filled with h2o that sank from the surface in the loftier latitudes . Warm water is more than buoyant than cold, which causes the upper sunlit layer to bladder on the denser deep ocean, with the transition between the two known as the "pycnocline" (for "density gradient") or "thermocline" (the vertical temperature gradient that drives density stratification across almost of the sea, Figure ii). Air current or another source of energy is required to bulldoze mixing across the pycnocline, and so the transport of water with its dissolved chemicals between the sunlit surface and the dark interior is sluggish. This dual issue of light on photosynthesis and seawater buoyancy is critical for the success of ocean phytoplankton. If the sea did not have a thin buoyant surface layer, mixing would acquit algae out of the light and thus away from their free energy source for most of the time. Instead of nearly neutrally buoyant single celled algae, larger, positively buoyant photosynthetic organisms (due east.g., pelagic seaweeds) might boss the open ocean. This hypothetical case aside, although feasible phytoplankton cells are found (admitting at low concentrations) in deeper waters, photosynthesis limits active phytoplankton growth to the upper skin of the ocean, while upper ocean density stratification prevents them from being mixed down into the dark abyss. Thus, most open ocean biomass, including phytoplankton, zooplankton, and nekton, is found within ~200 one thousand of the ocean surface.

Effigy 2
Typical weather in the subtropical sea, every bit indicated by data collected at the Bermuda Atlantic Time-series Station in July, 2008. The thermocline (vertical temperature slope) stratifies the upper h2o cavalcade. During this particular station occupation, the shallow wind-mixed surface layer is non well defined, presumably considering of stiff insolation and a lack of air current that allowed continuous stratification all the way to the surface. Very little sunlight penetrates deeper than ~100 k. New supply of the major nutrients Due north and P is express by the slow mixing across the upper thermocline (showing hither only the N nutrient nitrate, NO3 -). Within the upper euphotic zone, the slow nutrient supply is completely consumed by phytoplankton in their growth. This growth leads to the accumulation of particulate organic carbon in the surface bounding main, some of which is respired past bacteria, zooplankton, and other heterotrophs, and some of which is exported equally sinking material. The deep chlorophyll maximum (DCM) occurs at the contact where there is adequate light for photosynthesis and yet meaning food supply from beneath. The DCM should not be strictly interpreted as a depth maximum in phytoplankton biomass, as the phytoplankton at the DCM have a specially high internal chlorophyll concentration. The information shown here is fabricated available the Bermuda Plant of Bounding main Sciences (http://bats.bios.edu) and the Bermuda Bio Optics Project (http://www.icess.ucsb.edu/bbop/).
At the same time, the existence of a sparse buoyant surface layer conspires with other processes to impose nutrient limitation on ocean productivity. The export of organic affair to depth depletes the surface ocean of nutrients, causing the nutrients to accumulate in deep waters where there is no calorie-free available for photosynthesis (Figure 2). Because of the density difference between surface h2o and the deep ocean across virtually of the ocean, ocean circulation can only very slowly reintroduce dissolved nutrients to the euphotic zone. By driving nutrients out of the sunlit, buoyant surface waters, ocean productivity effectively limits itself.
Phytoplankton growth limitation has traditionally been interpreted in the context of Liebig'southward Law of the Minimum, which states that plant growth will be as swell as allowed by the to the lowest degree available resource, the "limiting nutrient" that sets the productivity of the organisation (de Baar 1994). While this view is powerful, interactions amidst nutrients and betwixt nutrients and lite can besides control productivity. A simple simply important example of this potential for "co-limitation" comes from polar regions, where oblique solar insolation combines with deep mixing of surface waters to yield low light availability. In such environments, higher iron supply can increase the efficiency with which phytoplankton capture light free energy (Maldonado et al. 1999, Sunda & Huntsman 1997). More broadly, it has been argued that phytoplankton should more often than not seek a state of co-limitation past all the chemicals they require, including the many trace metal nutrients (Morel 2008).
Who Are the Major Players in Sea Productivity?
Source: https://www.nature.com/scitable/knowledge/library/the-biological-productivity-of-the-ocean-70631104/
0 Response to "How Do You Know if an Element Is Limiting in Open Ocean? Graph"
Post a Comment